肿瘤免疫治疗小分子药物盘点 |
您所在的位置:网站首页 › pd1抗癌药的副作用 间质性肺炎 百度学术 › 肿瘤免疫治疗小分子药物盘点 |
肿瘤免疫治疗小分子药物盘点
|
前言
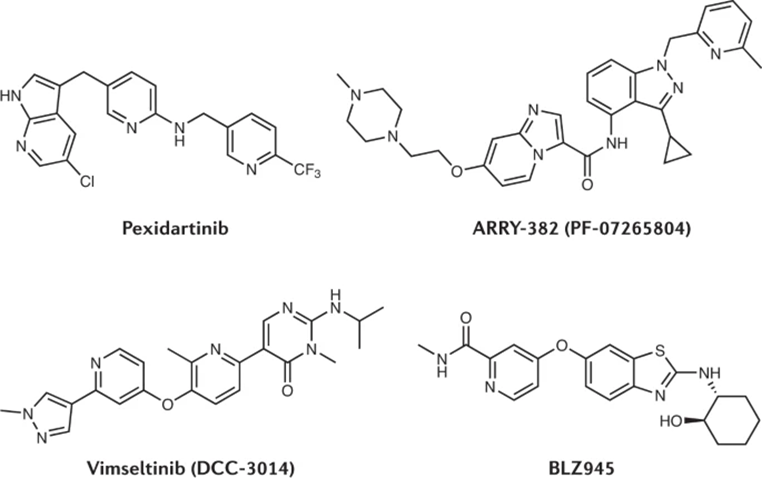
PD1、PDL1、CTLA4等免疫检查点蛋白抑制剂(ICIs)的成功, 使科学家们投入到肿瘤免疫治疗药物的开发热潮中。最近一些药物已进入临床试验,其中除了靶向表面蛋白的抗体外,不乏一些具有免疫刺激效应的小分子药物(见表1)。口服小分子药物克服了抗体药物不良反应多、半衰期长、输注给药不便等局限,为药物发现开辟了一个全新的领域。此外,肿瘤免疫小分子药物与免疫检查点抑制剂具有协同作用,两者联合用药的方案也得到了越来越多的批准。今年8月发表的Nature reviews盘点了肿瘤免疫治疗中的小分子药物及其应用发展 [1]。 表1:肿瘤免疫治疗部分小分子药物 靶向细胞生长和肿瘤代谢的小分子抑制类药物 01 靶向促肿瘤通路 癌症的两个固有的特征是失去细胞稳态周期控制和增加凋亡抵抗。另外,肿瘤持续的生长需要一个有利的微环境,并由促进血管生成、炎症和组织重塑的通路驱动。进展中的肿瘤能量需求不断增加,因此癌细胞重新调整其代谢途径,导致营养和代谢物浓度的显著变化,这些因素共同参与了肿瘤微环境(TME)的塑造 [2]。目前已经开发了各种靶向肿瘤细胞固有特征和TME途径的小分子药物。 1. 细胞生长抑制剂 细胞生长抑制类药物的疗效部分依赖抗肿瘤免疫反应的间接刺激,该类药物诱导的免疫原性细胞死亡(ICD)可以触发抗肿瘤T细胞应答的激活(图1)。在进入TME后,T细胞会遇到抑制信号而功能减弱,因此常常联合化疗和免疫检查点抗体。然而,化疗往往与淋巴减少症相关,会抵消诱导的适应性免疫反应。因此,联合具有选择性的靶向药物可能是更好的用药方案。 在BRAF(V600E)转移性黑色素瘤中,BRAF抑制剂与MEK抑制剂联合应用是治疗的标准方法,之后进展的患者常用PD-1抗体进行二线治疗。这使得研究人员探索BRAF/MEK抑制剂与ICIs的三联治疗。两项I期研究表明,BRAF/MEK抑制可导致ICD,产生促炎信号,增加CD8+T细胞浸润,提高MHC I表达水平,使肿瘤细胞对T细胞介导的攻击更敏感。在这种情况下,同时给予ICI会以协同方式维持T细胞活化。此外,小分子抗肿瘤药物的影响可能不止于诱导ICD。MEK抑制可在多个水平上直接调节免疫浸润的性质,例如通过减少CD4+调节性T细胞(Treg细胞)、髓源性抑制细胞(MDSC)和M2型肿瘤相关巨噬细胞(TAM),导致TME中CD8+/Treg淋巴细胞和M1/M2巨噬细胞比值增加。 一项II期疗效试验显示,与治疗组合达布雷尼布(BRAF抑制剂)和曲美替尼(MEK抑制剂)相比,加上帕博利珠单抗(PD1 抗体)的三联疗法延长了患者的无进展生存期。IMspire150 III期研究也也观察到同样的效果,这项研究评估了阿替利珠单抗(PDL1抗体)、维莫非尼(BRAF抑制剂)和考比替尼(MEK抑制剂)的联合,并于2020年7月获FDA批准用于治疗BRAF(V600E)突变型黑色素瘤。 图1:免疫原性细胞死亡调节T细胞活性 2. VEGF–VEGFR 抑制剂 血管内皮生长因子(VEGF)是一种刺激血管生成的信号蛋白,能抑制抗原提呈、招募免疫抑制细胞亚群、破坏抗肿瘤T细胞反应。高血管化肿瘤(如肾细胞癌)对VEGF–VEFGR通路抑制剂介导的血管生成阻断特别敏感。ICIs与这种药物的结合具有协同效益。2018年12月FDA首次批准贝伐单抗(VEGF 抗体)与阿替利珠单抗联合用于晚期非小细胞肺癌的一线治疗。2019年,阿西替尼成为首个FDA注册的小分子VEGFR酪氨酸激酶抑制剂,与帕博利珠单抗或阿维单抗(PDL1 抗体)联合用于晚期肾细胞癌的一线治疗。两种方案均优于迄今为止针对该适应症的标准方案——广谱激酶抑制剂舒尼替的单药治疗。 目前,在其他实体肿瘤(包括黑素瘤、头颈癌、肝细胞癌、结直肠癌、胃癌和妇科癌)中,已有超过100项类似方案的试验启动,总体结果有望获得FDA批准 [3] 。 3. 细胞因子、趋化因子通路阻断剂 细胞因子超家族在细胞交流中起着至关重要的作用,特别是在免疫系统中。干扰素-α (IFNα)和白细胞介素-2 (IL-2)对T细胞增殖和效应功能有强烈刺激作用,成为第一批用于癌症免疫治疗的细胞因子。然而,在大多数情况下,这些细胞因子治疗无法重现临床前模型的抗肿瘤疗效。迄今为止,阻断促癌炎症过程中的细胞因子通路和/或抑制T细胞免疫的治疗方法,如IL-1、IL-4、IL-6、IL-8、IL-10、TGFβ和CSF1,在临床研究中也没有表现出一致的疗效。尽管其中一些药物与ICIs和/或细胞抑制模式的组合可能被证明对癌症患者有效,但TME中细胞因子网络的复杂性使得单一通路抑制的抗癌效果削弱。 从免疫治疗的角度来看,抑制CSF1-CSF1R通路可能是一种更有前途的策略。高水平的CSF1会促进微环境中TAMs和MDSCs的浸润和扩张,在临床前模型中抑制该通路能减少髓细胞浸润并增加CD8+ T细胞,从而与ICIs协同作用。因此,该途径的多种抗体和小分子抑制剂或作为单一药物,或与细胞抑制药物或ICIs联合使用(图2)。 图2:CSF1R抑制剂结构 2019年,FDA批准小分子CSF1R-TK抑制剂培西达替尼用于治疗腱鞘巨细胞瘤 (TGCT)。ARRY-382是一种选择性更强的CSF1R抑制剂,在一项II期临床试验中与pembrolizumab联合治疗晚期实体肿瘤,但辉瑞公司最近发布的研究报告显示该方案对患者没有任何有益的效果。相比其他人类激酶,Vimseltinib对3类RTKs具有超过100倍的选择性。2022年9月12日,生物制药公司Deciphera Pharmaceuticals Inc.在ESMO上公布了Vimseltinib治疗腱鞘巨细胞瘤I/II期试验的最新数据,显示31名患者都表现出临床获益,A组的客观反应率为53%,B组为46%。另一种选择性CSF1R抑制剂BLZ945是一种脑穿透剂,目前正在进行I/II期临床试验,作为单药或与斯帕巴他珠单抗(PD1 抗体)联合用于治疗晚期实体肿瘤及中枢神经系统癌症。初步数据显示对胶质母细胞瘤患者具有抗肿瘤活性和可接受的安全性。 02 靶向代谢途径 肿瘤生长过程中各种代谢物的积累促进TME的形成,驱动这一过程的几种酶促蛋白可以被靶向调节,目前研究与小分子开发较为深入的主要为以下三种途径。 1. 谷氨酰胺和精氨酸代谢抑制剂 癌细胞通过促进糖酵解和谷氨酰胺分解来满足对能量和生物合成前体的高需求。值得注意的是,TME中不止有肿瘤细胞,T细胞、髓细胞以及其他基质细胞之间对葡萄糖、谷氨酰胺和其他营养物质存在激烈的竞争。尽管临床前模型中,用小分子抑制这些代谢通路可导致肿瘤生长减少和免疫细胞浸润重编程,但在临床试验中仍缺乏具体证据。如著名的口服谷氨酰胺酶抑制剂CB839,基于临床前的成功数据进行了多项临床研究。尽管如此,由于在两项针对肾细胞癌和非小细胞肺癌的试验中没有临床效益,Calithera Biosciences最近停止了该化合物的开发。 肿瘤、T细胞和MDSCs都依赖于精氨酸,从而进一步增加了这种氨基酸在TME中的需求。目前,一些小分子精氨酸酶抑制剂已经被开发出来,其中一种是硼酸基化合物INCB001158,已被Incyte纳入五项早期临床试验,在这些研究中,该药单独或与ICIs联合试验。初步试验报告表明,INCB001158的耐受性良好,临床疗效的信息有待全面评估发表。但截至最近,INCB001158已不再列在Incyte的投资组合中,这表明其临床开发已经停止。另外一种抑制剂是Calithera公司开发的CB-280,正在对囊性纤维化患者进行安全性评估。除上两者外,目前没有进一步的精氨酸酶抑制剂在临床试验中。 2. 腺苷途径阻断剂 腺苷是一种强大的免疫抑制剂,许多肿瘤产生高浓度的腺苷 [4]。其免疫抑制特性是由两个G蛋白偶联受体(GPCRs)介导的,即高亲和力的A2aR和低亲和力的A2bR(图3)。腺苷途径阻断剂imaradenant、ciforadenant和inupadenant特异性抑制A2aR,从2015年开始进行I/II期肿瘤试验,但迄今尚未报告临床疗效的明确证据。 目前科研人员的注意力转向双A2aR/A2bR抑制剂。一个例子是etrumadenant,两种受体的拮抗剂,其可联合zimberelimab(PD1 抗体)和多西他赛(多烯紫杉醇)治疗转移性去势抵抗性前列腺癌患者。此外,在结直肠癌中,etrumadenant联合FOLFOX化疗作为三线治疗显示出有临床意义的疗效。另外,考虑到TME中的高腺苷浓度,高亲和力A2aR可能会被永久打开,其他研究人员正在开发低亲和度A2bR的阻断剂。Teon Therapeutics已开始开发TT-702,这是一种选择性A2bR拮抗剂,对免疫细胞和癌细胞都有活性。Taros Therapeutics也在开发A2bR选择性分子。 在TME中,腺苷由外胚层酶CD73催化AMP生成,因此CD73成为该途径另一重要靶点。除了几种Ab类药物外,还有AB680和OP5244/ORIC-533两个小分子正在研究。此外,腺苷可以通过各种额外的外泌酶产生,如CD38、CD39、CD203a/外泌核苷酸磷酸磷酸酶/磷酸二酯酶家族成员1、碱性磷酸酶和前列腺酸性肽酶(图3)。目前,针对CD38和CD39的临床抗体已经存在,未来可能会产生更多针对这组外胚层酶的小分子药物。该途径之后的方向可以聚焦在正确靶点组合的选择上。 图3:腺苷途径相关通路 3. 犬尿氨酸途径抑制剂 肿瘤细胞和间质细胞可以在IDO1和TDO两种酶介导的反应中分解色氨酸,从而产生犬尿氨酸 [5]。由此导致的色氨酸缺乏和TME中犬尿氨酸的积累都会抑制免疫反应。在与芳香烃受体(AHR)结合后,犬尿氨酸可以抑制效应T细胞,促进 CD4+ Treg细胞和MDCSs的发展(图4)。临床实验表明,该途径的阻断在IDO1和犬尿氨酸水平高的患者中更具价值。林罗多司他是一种更有效的、不可逆的IDO1抑制剂, 耐受性良好,目前正与纳武单抗联合用于黑色素瘤患者的III期临床试验中。 但是,其他色氨酸降解酶TDO和IDO2的代偿性表达可能使肿瘤对IDO1抑制不敏感。此外,IL-4诱导-1(IL4i1)最近被确认为一种额外的色氨酸分解酶,由肿瘤和髓系细胞分泌到TME(图4)。因此,可能需要同时抑制上述几个靶点,从而提高临床试验中的疗效。最近已报道IDO1/TDO双抑制剂(如RG-70099和M4112)。另外,两种AHR阻断剂(BAY2416964和IK175)已进入实体肿瘤的临床试验。 图4:犬尿氨酸途径介导的抗肿瘤免疫 靶向固有免疫和免疫检查点的小分子药物 03 靶向固有免疫通路 最早时曾在肿瘤内注射Coley疫苗和卡介苗(BCG)治疗癌症,尽管当时并不清楚作用机制,但现在我们知道,注射细菌提取物会触发多种固有免疫途径,从而介导肿瘤细胞的杀伤。 1. TLR 激动剂 Toll样受体(TLR)是一个1型跨膜蛋白家族,通过触发固有反应和介导适应性反应的激活,在协调病原体免疫方面起着关键作用 [2]。病原体相关分子模式(PAMPs)作为TLR配体,促进新表位的交叉呈递,在引流淋巴组织中激活树突状细胞。到目前为止,三种TLR激动剂已被批准用于肿瘤免疫治疗:用于膀胱癌局部免疫治疗的TLR2/TLR4刺激物卡介苗、用于局部治疗选定上皮病变的小分子TLR7/TLR8配体咪喹莫特,以及单磷酸脂A(MPL),一种TLR4配体,用作预防性人乳头瘤病毒(HPV)疫苗Cervarix95的佐剂。 许多TLR激动剂已经在临床试验中进行了研究,其中大多数靶向内质体TLR。尽管在临床前肿瘤模型中使用各种TLR激动剂是有效的,但临床试验的结果一直令人失望,这主要有三个主要原因。一是TLR在许多不同的细胞类型上表达,包括不同程度上的肿瘤细胞,使得TLR激动剂的体内作用机制难以预测;二是TLR的表达模式和参与的生物学反应在小鼠和人类之间有很大差异;三是TLR激动剂全身给药有诱导全身免疫激活的高风险,与细胞因子释放综合征等严重不良事件相关。 但TLR7激动剂LHC165克服了一些困难。它被吸附到氢氧化铝上,使其从注射部位缓慢释放,激活局部受限的免疫。据报道,在晚期实体瘤患者中首次出现临床抗肿瘤活性,包括作为单药或与sparvatigumab(PD1抗体)联合使用,尤其是对于基线时免疫微环境活跃的患者。为了降低全身给药时的毒性风险,目前正在探索通过抗体-药物结合物(ADC)靶向给予TLR激动剂。例如,SBT6050将强效TLR8激动剂与抗HER2抗体结合起来治疗HER2表达实体肿瘤。 2. STING激动剂 除TLR外,其他“危险”传感器主要响应细胞内源性分子信号(DAMPs),从而为激活先天免疫和适应性免疫提供额外途径。干扰素基因刺激物(STING)通路对非凋亡细胞死亡释放的DNA片段作出反应。这些DNA片段与细胞溶质酶cGMP–AMP合成酶(cGAS)的结合,导致环状二核苷酸cGAMP的产生,并激活内质网相关蛋白STING。随后,STING与TANK结合激酶1(TBK1)结合,然后该复合物转移到核周区域并激活下游介质,特别是转录因子干扰素调节因子3(IRF3)(图1)[3]。 临床上第一代STING激动剂主要基于环状二核苷酸(CDN),如ADU-S100和MK-1454。但这两种化合物远远落后于预期,临床开发已被终止。与TLR激动剂类似,这些化合物的一个关键限制是需要肿瘤内注射。第一种全身给药的STING激动剂SB 11285和SNX281正在与PD1/PDL1抗体联合进行临床评估。此外,含有氨基苯并咪唑框架的非CDN基STING拮抗剂在小鼠中的药代动力学和全身活性得到改善。GSK3745417作为单一疗法以及与帕博利珠单抗联合治疗晚期实体瘤正在进行I期临床试验。 当然,STING激活仍存在一些不确定性。过度刺激cGAS–STING途径有诱发细胞因子风暴的风险,最近的临床前数据表明,STING激活可通过触发应激和促凋亡途径诱导T细胞死亡。 3. ENPP1、TREX1和PARP7抑制剂 鉴于直接激活STING所面临的挑战,最近的策略侧重于以间接方式刺激该通路,如针对STING负调控因子ENPP1。ENPP1降解免疫刺激代谢物cGAMP和ATP,从而参与免疫抑制腺苷的生成(图1)。抑制ENPP1可通过增加cGAMP水平导致STING激活,同时减少对腺苷通路的刺激。一些口服ENPP1抑制剂已经进行了临床前研究,如MV626、SR-8314及其后续分子SR-8541。到目前为止还没有开始临床试验。 进一步激活STING途径的方法是抑制3 ' 核酸修复外切酶1 (TREX1),它负责降解肿瘤来源的DNA。抑制TREX1会导致胞质DNA的积累和STING通路的激活。小分子TREX1抑制剂的首批实例最近由Constellation Pharmaceutical和Venenum Biodesign LLC公布。 多聚ADP核糖聚合酶7(PARP7)在吸烟相关性强的癌症中过度表达,通过与TBK1相互作用负调节I型干扰素反应。因此,它也作为STING途径的抑制剂(图1)。RBN-2397是该酶的一种有效特异性阻断剂,最近已进入I期安全试验。 图1:STING相关通路 04 小分子免疫检查点抑制剂(ICIs) 尽管免疫检查点抗体已成为免疫治疗的金标准,但小分子药物相对于抗体药物的潜在优势引发了干扰PDL1–PD1轴的药物化学研究。另外,随着对T细胞内信号的不断深入研究,发现了T细胞受体(TCR)参与下游的几个负反馈回路,这些回路可能增强抗肿瘤T细胞免疫(图2)。主要的成药靶点包括造血祖激酶1(HPK1或MAP4K1)、二酰基甘油激酶(DGKα和DGKζ)、酪氨酸蛋白磷酸酶非受体型(PTPN6和 PTPN22)、E3泛素蛋白连接酶CBL-B等。 图2:参与免疫受体信号传递的胞内靶点 1. PDL1–PD1小分子抑制剂 长期以来,PDL1-PD1的相互作用被认为不受小分子的抑制。尽管如此,第一批口服小分子药物CA-170(图3)和GS-4224已进入临床。CA-170来源于氨基酸丝氨酸-天冬酰胺-苏氨酸序列,该序列是由PD1初级序列的基序得到的。据报道,该化合物可靶向PDL1和VISTA。VISTA是一种检查点调节剂,在髓细胞上高度表达。核磁共振光谱显示CA-170在不阻止络合物形成的情况下产生缺陷的PD1-PDL1三元络合物,且在小鼠体内的口服生物利用度为50%。而GS-4224的安全性研究已被终止。 另一种分子ARB-272572(图3)在亚纳摩尔浓度下可干扰PDL1功能,并在可移植的同源结直肠癌小鼠模型和肝炎病毒感染模型中显示出活性。一种相关的联苯分子INCB086550(图3)正在实体肿瘤患者中进行I/II期临床研究。口服这种化合物可抑制过表达PDL1的MC38肿瘤,其效力与阿替利珠单抗相当。这些化合物口服似乎增加了T细胞肿瘤浸润,不仅通过与PDL1结合,还通过诱导PD1的细胞内化。第一批小分子ICIs进入临床是十分重要的一步,但这些化合物在哪些方面可以达到临床上抗体的最佳状态,仍有待观察。 图3:检查点小分子抑制剂 2. MAP4K1抑制剂 MAP4K1是一种丝氨酸/苏氨酸激酶,仅在造血细胞类型中表达 [4]。通过其富含脯氨酸的结构域,MAP4K1能够与造血细胞中的各种配体结合,MAP4K1的功能已经在TCR信号环境中得到了详细研究。TCR刺激后,MAP4K1在酪氨酸381上磷酸化,随后,MAP4K1被募集到TCR信号复合体中,通过其丝氨酸/苏氨酸激酶功能诱导该复合体解离,起到负调控作用。 最近,一些论文揭示MAP4K1是小分子干预的合适靶点。最早发表的MAP4K1抑制剂之一是ZYF0033。据报道,该化合物可以抑制MAP4K1,影响近端生物标志物pSLP76,在4T1同基因小鼠模型中剂量依赖性的抑制肿瘤生长。BAY-405是另一个最近公布的MAP4K1抑制剂。该分子可以抑制SLP76的磷酸化,具有亚微摩尔的效力,并显示出良好的激酶选择性。体外功能分析显示BAY-405可增强人细胞毒性T细胞中细胞因子的活化和产生。BAY-405在同基因EMT6乳腺和B16-OVA黑色素瘤小鼠模型中显示了单药疗效,在B16-OVA模型中与抗PDL1 Ab联合使用时也有疗效。 一些MAP4K1抑制剂已在实体肿瘤中进行了试验,例如NDI-101150/NMBS-2,具有很强的细胞效力,对所有MAP4K家族成员具有>100×的选择性。NDI-101150随后在一组同基因体内疗效模型中进行了评估,12个模型中有4个被观察到单药治疗肿瘤生长减少。重要的是,在EMT6同基因乳腺肿瘤模型中观察到肿瘤完全减少。联合抗PD1 Ab可以恢复衰竭T细胞的细胞因子分泌,并在CT26肿瘤模型中抑制肿瘤生长。此外,MAP4K1抑制剂CFI-402411的I/II期试验已开始评估单药和与帕博利珠单抗联合使用的耐受性。另外三种MAP4K1小分子抑制剂BGB-15025、PRJ1-3024和PF-07265028也已进入临床研究。 3. DGKα和DGKζ抑制剂 二酰基甘油(DAG)是磷脂酶Cγ1在TCR作用时产生的第二信使,触发信号级联,在T细胞发育和功能中起重要作用。DGKs家族催化DAG磷酸化为磷脂酸,从而调节DAG介导的信号。其中,DGKα和DGKζ在T细胞中显著表达 。两种DGKs的表达和活性在TCR刺激下增强,而持续表达与肿瘤浸润T细胞的低反应性相关(图2)。 DGKα小分子抑制剂R59022, R59949和利坦色林已经被发现几十年。这些第一代DGK抑制剂的体外实验已证明通过药理抑制DGKs增强T细胞活性的概念,以及DGK抑制与PD1 阻断的联合作用。在小鼠中,用利坦色林治疗可以克服对PD1阻断的耐药性。百时美施贵宝最近公布的一项专利申请声称双DGKα/DGKζ抑制剂是T细胞激活剂。其中一种抑制剂对DGKα的抑制作用低于1毫摩尔。目前开发临床DGKs抑制剂的一个关键顾虑是脱靶效应。 4. CBL-B抑制剂 泛素化,即泛素部分附着在蛋白质赖氨酸残基上,是蛋白质的翻译后修饰,参与调节多种信号通路和细胞过程。CBL-B是一个RING家族E3泛素连接酶,作为TCR激活的负调控下游发挥作用 [5] 。其活性被CD28共刺激抑制,并被PD1和CTLA4接合诱导。 最初的CBL-B靶向研究集中在非小分子方法上,包括干扰CBL-B功能的细胞渗透性肽和通过小干扰RNA调控CBLB基因表达或靶向基因组编辑。2016年,Progenra提出了一系列已知的小分子CBL-B抑制剂,能够通过抑制TAM受体的泛素化激活T细胞和NK细胞活性,但缺乏后续研究。目前已知的唯一CBL-B抑制剂,由Nurix开发,可增强体外T细胞的反应性,以及可移植的同基因小鼠模型中T细胞依赖性肿瘤的消退。化合物NX-1607在晚期恶性肿瘤中的安全性试验最近已经启动。泛素系统的复杂性和对底物特异性的未知提高了选择性小分子CBL-B抑制剂的发现门槛。 5. 磷酸酶 几种磷酸酶(包括SHP1、SHP2和PTPN22等)介导T细胞信号转导的负调控。SHP099是一种选择性的、口服的SHP2抑制剂,在免疫缺陷小鼠中显示了对人类肿瘤异种移植的抗肿瘤效果。另一种SHP2抑制剂RMC-4550的试验显示,其在CT26和EMT6同基因肿瘤模型中具有中等的抗肿瘤效果,同时对PDL1和CTLA4抗体也具有补充作用。几种SHP2抑制剂正在与ICI Abs联合进行临床评估。TNO155与斯巴达珠单抗联合、RMC-4630与帕博利珠单抗联合都处于I期临床试验中。 结 语 胞生长抑制剂、VEGF–VEGFR 抑制剂和细胞因子、趋化因子抑制剂等传统小分子药与ICIs联合应用具有更大的潜力。针对代谢途径的阻断剂可以改变肿瘤或TME中其他免疫细胞的行为,从而激活对肿瘤的杀伤反应。 科研人员正在全力扩大肿瘤免疫治疗的方案,其中小分子药物发挥着越来越重要的作用(图4)。尽管一些靶向促肿瘤途径的小分子已被批准作为PD1-PDL1抗体的伴侣,但未来几年将探索小分子是否也可以应用于临床,直接刺激T细胞和/或先天抗肿瘤免疫。 图4:TME造成的免疫调节障碍与解决者小分子 参考资料: [1] Offringa R, Kötzner L, Huck B, Urbahns K. The expanding role for small molecules in immuno-oncology [published online ahead of print, 2022 Aug 18]. Nat Rev Drug Discov. 2022;10.1038/s41573-022-00538-9. doi:10.1038/s41573-022-00538-9. [2] Hanahan D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022;12(1):31-46. doi:10.1158/2159-8290.CD-21-1059. [3] Hack SP, Zhu AX, Wang Y. Augmenting Anticancer Immunity Through Combined Targeting of Angiogenic and PD-1/PD-L1 Pathways: Challenges and Opportunities. Front Immunol. 2020;11:598877. Published 2020 Nov 5. doi:10.3389/fimmu.2020.598877. [4] Allard B, Allard D, Buisseret L, Stagg J. The adenosine pathway in immuno-oncology [published correction appears in Nat Rev Clin Oncol. 2020 Jul 17;:]. Nat Rev Clin Oncol. 2020;17(10):611-629. doi:10.1038/s41571-020-0382-2. [5] Opitz CA, Somarribas Patterson LF, Mohapatra SR, et al. The therapeutic potential of targeting tryptophan catabolism in cancer. Br J Cancer. 2020;122(1):30-44. doi:10.1038/s41416-019-0664-6. 作者 | 梨子君 |









【本文地址】
今日新闻 |
推荐新闻 |